地球外生命は存在するのか?
この究極ともいえる問いを
「原始生命の発生確率」そして
「最新の宇宙研究の成果」
をもとに考察していく話題の新刊
『宇宙になぜ、生命があるのか』。
本書では、
宇宙物理学者である著者が、
有機物からRNA、DNAの合成、
そして原始生命の誕生について考えながら、
138億年のわれわれの宇宙において、
現在考えられているさまざまな理論をもとに、
それが「奇跡」だったのかを検証していきます。
今回は、この本の中から、
生命という現象を物理学の視点から
見るとなにが見えるのか、
考えていきます。
DNAの合成はエントロピーの
法則に反するのか?
ここで、
生命活動の根幹をなす現象を物理学の
観点からあらためて見直してみることにしたい。
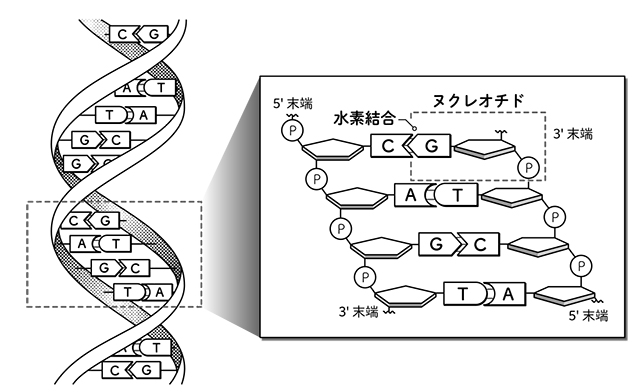
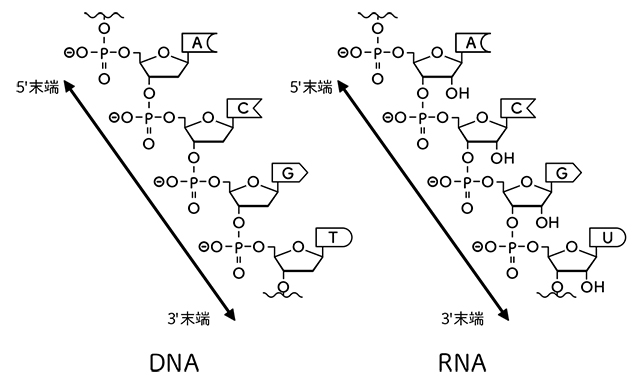
DNAもタンパク質も一本の鎖であり、
その鎖をつなげていくには外から
エネルギーを与えることが必要である。
外からエネルギーが与えられて、
バラバラだったアミノ酸やヌクレオチドが、
秩序を持った一本鎖に組み上がるというのは、
やはり熱力学的には奇異に感じる。
普通は、
例えば陽子と電子が結合した水素原子でも、
酸素原子と水素原子が結合した水分子でも、
バラバラなものが結合する場合は
それらの間に引力が働き、
余分なエネルギーが生じて
それを外に捨てるものである。
地上で重力に任せて物を落とせば、
引力によって物体の運動エネルギーが
生じることを思い起こせばいい。
物が地面に激突すれば、
運動エネルギーは熱エネルギーに転化し、
それは周囲に散逸してしまう。
これがエントロピーの増大で、
この逆は自然には起こらない。
ところがDNAやタンパク質の合成はその逆で、
結合する際にエネルギーを吸うのである。
この一見、
自然法則とは逆に見える反応が
起こる秘訣は何なのか。
DNAやたんぱく質の合成を見ていくと
それは、
局所的にはエントロピーが
減少しているように見えても、
より大局的に見てエントロピーが
増大していれば、
熱力学の法則に反しないということである。
とはいえ、
この説明では抽象的すぎて
納得できない向きもあろう。
そこで具体的に、
DNAやタンパク質の合成反応を吟味しつつ、
どうして全体としてはエントロピーが
増大しているといえるのか、
を見ていきたい。
エアコンで冷却する例を思い出すと、
局所的にエントロピーが減少して
冷却が起こる秘訣は、
全体として見れば外からエネルギーが供給され、
そのエネルギーが最終的には
外界に散逸することであった。
この、
「外界に散逸」する過程により
エントロピーが増大し、
逆方向、
つまり外に散らばってしまった
熱エネルギーが自然にエアコンに
戻ってくるようなことは起こらない。
DNAの複製の過程でも、
本質的に同じことが起きている。
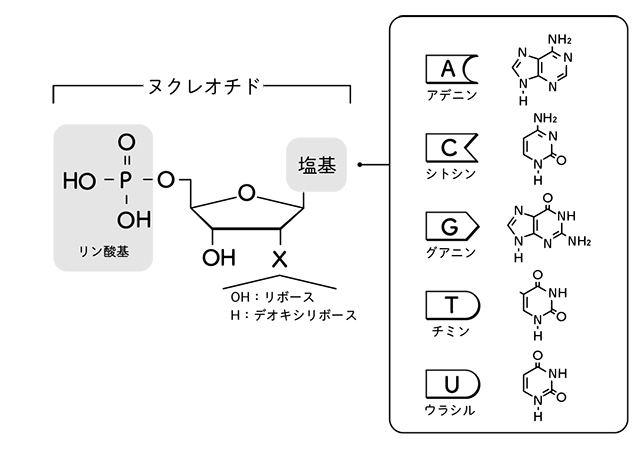
ヌクレオチドを結合してDNAの
一本鎖にするには
外からのエネルギーが必要で、
そのエネルギー源はヌクレオチドについている
高エネルギーリン酸結合である。
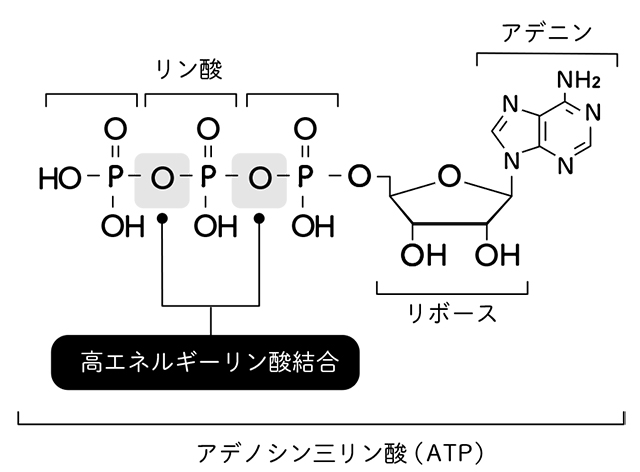
DNAとして結合する前の個体のヌクレオチドは、
3つのリン酸がつながったもの
(ヌクレオシド三リン酸)である。
塩基がAのものがまさにATP、
つまり生物のエネルギー通貨だ。
この、
リン酸が連なった部分に
エネルギーがたっぷりと
蓄えられている。
そしてDNA合成酵素によって、
作製中のDNAにヌクレオチドが結合される際に、
このうち2つのリン酸がとれて、
DNA中のヌクレオチドのリン酸は1つだけになる。
このリン酸結合が切断される際に
大きなエネルギーが解放され、
ヌクレオチドの結合に使われる。
そして2つのリン酸が連なったリン酸基は
周囲に捨てられる。
この、
リン酸基が周囲に放出される過程が
実に重要である。
リン酸結合の切断で解放されたエネルギーは
ヌクレオチドの結合だけでなく、
捨てられたリン酸基にも
運動エネルギーとして与えられ、
それはすぐに周囲の分子と
相互作用する過程で散逸してしまうであろう。
これこそ、
バラバラのヌクレオチドが
一本鎖につながるという、
エントロピーが減少して見えるような
反応が進行するからくりである。
本当にエントロピーが増大しているのか
本当にエントロピーが増大しているか
どうかを吟味するには、
その反応の逆の過程が起こりうるか、
を考えればよい。
捨てられたリン酸基の運動エネルギーが
散逸する前ならば、
そのエネルギーを保持したリン酸基を
DNAにぶつけて、ヌクレオチドを切り離し、
3つのリン酸が結合した元の
単体ヌクレオチドに戻す反応が可能である。
だが実際は、DNA
の周囲にあるリン酸基は、
捨てられたときに持っていたエネルギーを
すぐに周囲に散逸してしまっており、
この逆反応を引き起こすためのエネルギーを
獲得する手段がない。
このためDNAを分解する
方向の反応は起こらず、
一方、
リン酸が3つ結合したヌクレオチドさえ
豊富にあれば、
リン酸結合のエネルギーを使ってDNAの
合成反応はどんどん進むことになる。
リン酸結合に蓄えられたエネルギーが、
一部はDNAの合成に使われるが、
残りは外界に捨てられて散逸していることが、
DNA合成を推し進める駆動力なのである。
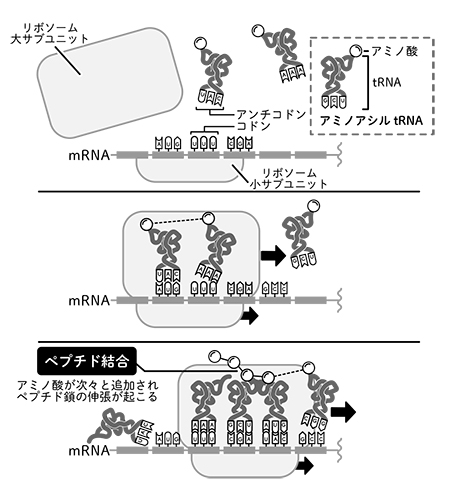
タンパク質の合成でも同様である。
アミノ酸を結合してタンパク質をつくるためにはまず、
アミノ酸とtRNAを結合し、
アミノアシルtRNAとして
「活性化」させる必要があった。
この際に、
エネルギー通貨ATPが用いられる。
つまり、
アミノアシルtRNAはエネルギーを
豊富に蓄えた状態である。
リボソームでは、
これをtRNAとアミノ酸に分解する際に
解放されるエネルギーを用いて、
アミノ酸を合成中のタンパク質に結合させる。
同時に、
不要となったtRNAは捨てられる。
アミノアシルtRNAのエネルギーの一部が、
捨てられたtRNAを通じて周囲に散逸することこそ、
反応をタンパク質の分解ではなく
合成の方向に進める原動力であることは、
もう読者にはおわかりであろう。
では、
ヌクレオチドのリン酸結合や
アミノアシルtRNAに蓄えられたエネルギーは
どこからくるのか。
それは大元をたどれば、
生体の外から供給されたエネルギー
(植物なら光合成のための光、
動物なら捕食した生物の有機物)である。
一見、
エントロピーが減少するかのように見える
DNAやタンパク質の合成こそ、
生命の秩序や構造を生み出す根源の力であるが、
それはあくまで、
物理学の法則に違うことなく
粛々と反応が進んでいるのである。
<参考:文= 戸谷 友則>
1喧嘩はするな、
2意地悪はするな、
3過去をくよくよするな、
4先を見通して暮らせよ、
5困っている人を助けよ、
あなたなら出来ます応援しています